Developmental Milestones
- Developmental Cell paper
- Video Abstract
- Preview by Michael K Richardson: “A Phyloptypic Stage for All Animals?”
- Full Datasets: At GEO: GSE31422, GPL14142, GPL14143, GPL14144, GPL14145, and GPL14146
The complex process by which an animal develops from a single cell to a multi-cellular differentiated organism has been organized by biologists into morphological stages. The appearance of these stages may hint at the modularity of embryonic development, or rather, that “stages exist in the mind of the biologist, and not in the larva” (Hickman, 1999) or embryo. It thus remains unclear whether development at the gene regulatory level reflects the stage-like processes occurring morphologically.
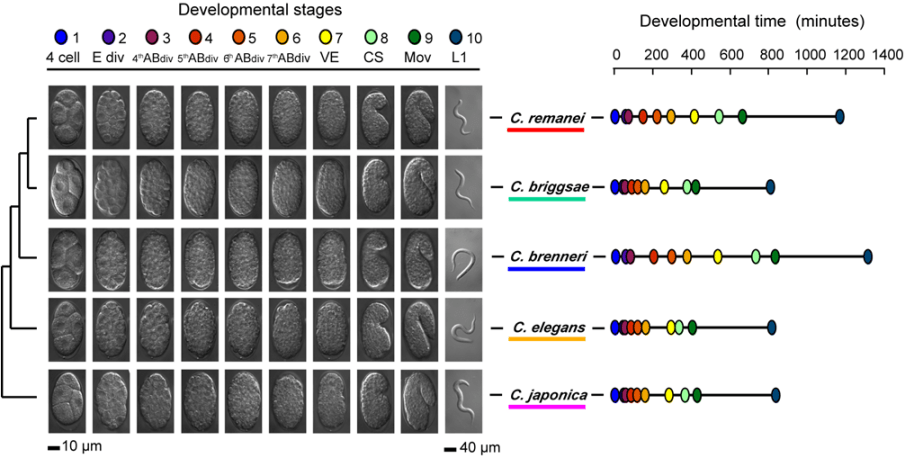
Development may be monitored through gene expression analyses with mRNA levels constituting an efficient proxy for the gene regulatory states underlying embryonic processes. To examine the relationship between morphological stages and their underlying molecular activity, we compared the transcriptomes of five nematode species developing at different rates, with the assumption that when the relationship is present transcriptomes at a developmental stage will be similar across species regardless of the time between them. We chose ten nematode developmental stages ranging from the 4-cell stage which comprises mostly maternal transcripts to the free-living first juvenile (L1) stage. Embryonic timings of each developmental stage vary with respect to C. elegans on average by 22% across the other four species (see figure).
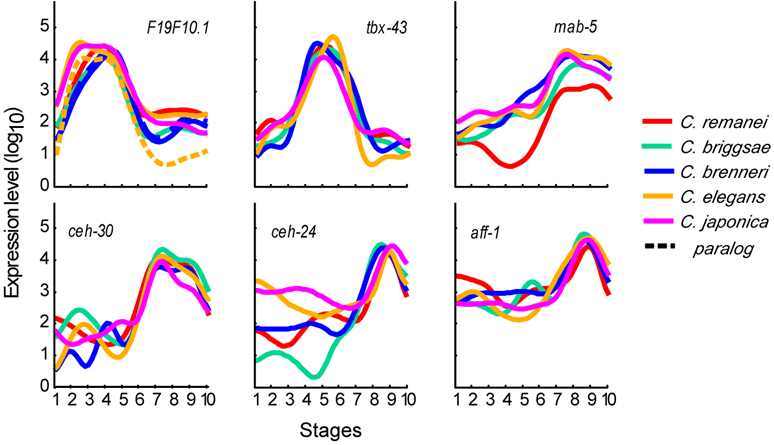
 The resulting transcriptomes reveal dynamic gene expression profiles across development. For example, 6,790 C. elegans genes were found to be dynamically expressed across the 10 sampled stages. Shown are gene expression profiles for six orthologous groups across the developmental stages.
The resulting transcriptomes reveal dynamic gene expression profiles across development. For example, 6,790 C. elegans genes were found to be dynamically expressed across the 10 sampled stages. Shown are gene expression profiles for six orthologous groups across the developmental stages.
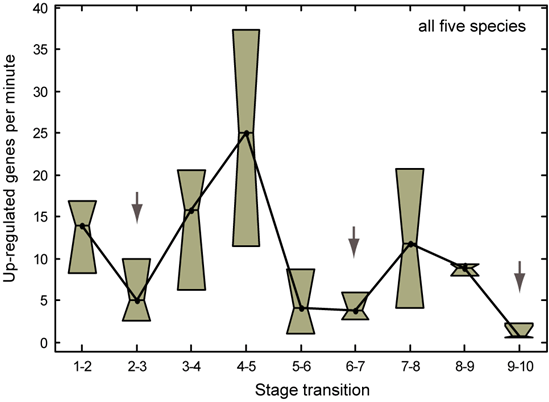
 We initially examined the dynamics of gene expression across stages to ask whether expression turnover is continuous across developmental time in C. elegans as previously reported (Baugh et al., 2003) or if the dynamics depend upon time (Bozinovic et al., 2011). To determine this, we computed the number of genes significantly up-regulated between adjacent stages and normalized by the number of intermittent minutes. We found an uneven pattern of expression change among the C. elegans stages. We next repeated this analysis in the other four species and found a similar oscillatory pattern. Interestingly, we found that entry into stages 3 (gastrulation) and 7 (ventral enclosure) is followed by a more than two-fold acceleration in the number of expressed genes per minute. These “bursts” of expression suggest that stages 3 and 7 are unique in terms of the number of transcripts being up-regulated per minute during their transitions relative to the stages that follow.
We initially examined the dynamics of gene expression across stages to ask whether expression turnover is continuous across developmental time in C. elegans as previously reported (Baugh et al., 2003) or if the dynamics depend upon time (Bozinovic et al., 2011). To determine this, we computed the number of genes significantly up-regulated between adjacent stages and normalized by the number of intermittent minutes. We found an uneven pattern of expression change among the C. elegans stages. We next repeated this analysis in the other four species and found a similar oscillatory pattern. Interestingly, we found that entry into stages 3 (gastrulation) and 7 (ventral enclosure) is followed by a more than two-fold acceleration in the number of expressed genes per minute. These “bursts” of expression suggest that stages 3 and 7 are unique in terms of the number of transcripts being up-regulated per minute during their transitions relative to the stages that follow.
As a second metric for change in gene expression throughout development, we computed for each pair of stages the expression ‘divergence’, defined as unity minus the correlation coefficient between their transcriptomes (1-R). A heatmap of such divergences for the C. elegansdevelopmental transcriptome is shown in the figure below. This map reveals that adjacent stages are more similar than temporally distant stages, reflecting the progression of development. The map also defines four clusters of stages within which transcriptome variation is low: the maternal transcriptome, the gastrulation transcriptome, the morphogenesis transcriptome, and the juvenile transcriptome.
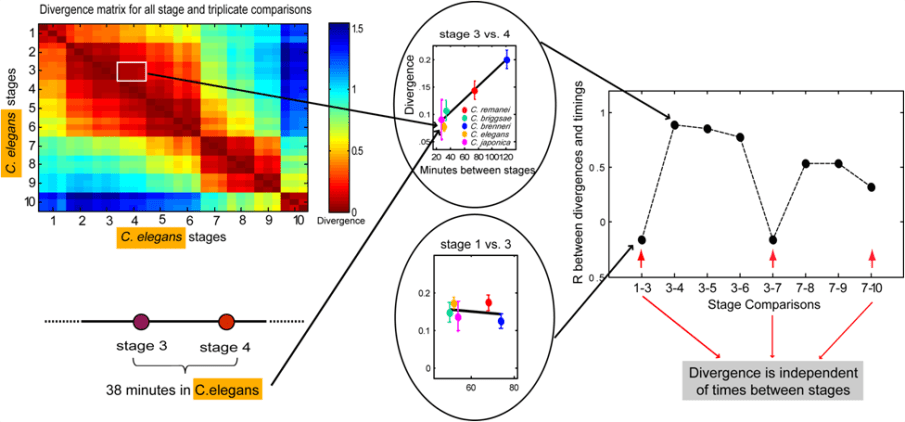
The divergence metric allowed us to ask whether pairs of stages show a correlation between elapsed time and the transcriptomic change between them, across the same stage pairs for all five species. For example, between stages 3 and 4 there are 130 minutes in C. brenneri, but only 38 minutes in C. elegans. Correspondingly, we found more transcriptome divergence in C. brenneri than in C. japonica between these stages. Thus, there is a time-dependent transcriptomic divergence between stages 3 and 4.
In applying this analysis to stages 3 and 7, which we previously found to exhibit unique characteristics in their expression dynamics, we discovered that these stages also exhibit time-independence with respect to their expression divergence. Intermediate stages show a correlation between time and divergence, but this correlation is lost upon entry into stages 3 and 7. We therefore concluded that these stages meet our time-independence criterion and should be viewed as developmental milestones, while intermediate stages may be seen as transitions.
To examine the transcriptome data at the functional level, we classified each gene to one of nine temporal expression classes depending upon the stage of activation, and queried for enrichment of diverse functional sets of. The resulting enrichments for the functional classes provide a map of the modules of genes up-regulated during development. Our enrichment analysis confirmed that genes that are activated towards stage 7 are disproportionally enriched with crucial functions such as key developmental processes of tissue differentiation (muscle and neuron), protein domains such as homeobox domains, C2H2-type zinc fingers, immunoglobulin-like domains, collagens, SH3 domains, PDZ and PH domains (involved in cell-cell signaling) and significant biological functions such as cell-cell interactions, morphogenesis and locomotion. The enrichment of homeobox transcription factors – known regulators of anterior-posterior pattern formation – is particularly significant (P<10-12, hypergeometric test). These functional enrichments suggest that the ventral enclosure milestone (stage 7) may serve as the phylotypic stage of these species, which has proved elusive in nematodes.
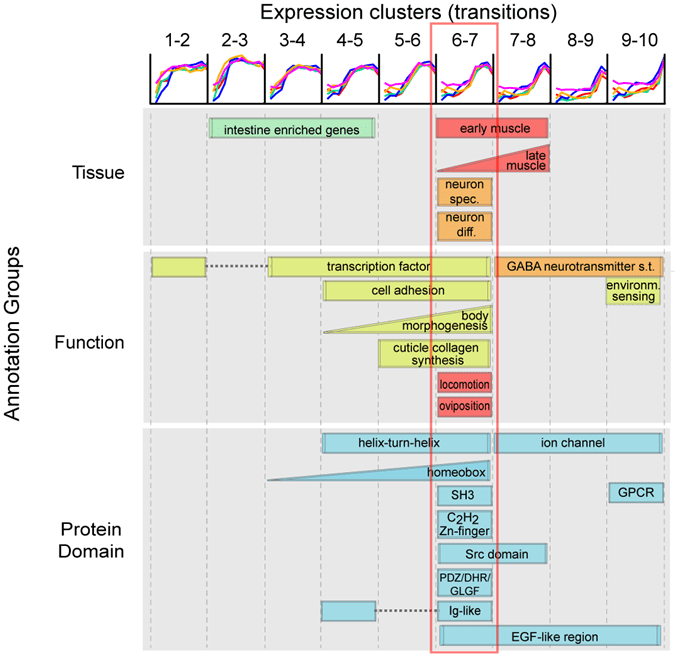
The phylotypic stage is a stage in mid-embryogenesis where divergent species appear most similar (Irie and Kuratani, 2011; Kalinka et al., 2010; Raff, 1996). Ventral enclosure (stage 7) has already been established as a period of intense integration of cell types (Chin-Sang and Chisholm, 2000). Our analysis further identifies a lack of a correlation at this stage between expression divergences and timing changes across species where the relative lull in expression is enriched with the up-regulation of homeobox transcription factors, characteristic of the phylotypic stage in other phyla (Slack et al., 1993).
The phylotypic stage is important as it suggests that evolutionary constraints are stronger for this stage than in earlier or later stages (Raff, 1996). Thus, if stage 7 indeed serves as the nematode phylotypic stage then the genes expressed in this stage may be enriched for knock-down phenotypes that reflect their importance. To test this, we searched for enrichments of all phenotypes found by RNAi screenings (Kamath et al., 2003) across the nine temporal expression classes. Strikingly, we found that stage 7 was uniquely enriched with perturbation phenotypes including dumpy, locomotion abnormal, morphology abnormal, and paralysis. This enrichment pattern is consistent with the notion that the ventral enclosure stage is the nematode phylotypic stage.
Collectively, our results provide five lines of evidence for a nematode phylotypic stage occurring during ventral enclosure (stage 7): 1. a constriction of expression divergence, also observed in the phylotypic stages of arthropods and chordates (Irie and Kuratani, 2011; Kalinka et al., 2010), 2. expression of crucial developmental regulators during this stage, 3. enrichment of RNAi phenotypes during this stage, 4. A subsequent burst of expression, and 5. time-invariance with respect to expression divergence across stages.
To test whether this nematode transcriptional signature is conserved across more distant phyla and is perhaps a general characteristic of Bilaterian embryogenesis, we examined the embryonic transcriptome of the amphibian Xenopus tropicalis. A recent X. tropicalis dataset(Yanai et al., 2011) was designed using the same probe selection strategy, platform, and internal controls, allowing for a meaningful comparison with our C. elegans dataset. We examined genes that were up-regulated in X. tropicalis during the gastrulation, neurulation, post-neurulation, tail-bud, and post-tail-bud stages. We assigned each dynamically expressed gene to a particular stage, delineated orthologs across the two species, and computed the statistical enrichment of the overlap of the orthologs in each pair of C. elegans and X. tropicalisstages. The resulting pattern of enrichments shows that C. elegans stages preceding stage 7 are not significantly conserved in any X. tropicalis stage, including gastrulation. Starting, however, at ventral enclosure (stage 7), C. elegans up-regulated genes are enriched with genes expressed during the X. tropicalis tail-bud stage, the chordate phylotypic stage (Raff, 1996). This significant overlap suggests that at the transcriptional level, the phylotypic stage is conserved across the Bilateria, as has been previously suggested (Domazet-Loso and Tautz, 2010; Kalinka et al., 2010; Slack et al., 1993). These genes are enriched with functions in locomotion, transcriptional activity, in particular helix-loop-helix, and hedgehog signaling. Expression of muscle-specific genes is also enriched among this gene set (P<10-14), functionally uniting the previous enrichments with their known role in this tissue (Ingham et al., 2011; Massari and Murre, 2000).
Another significant enrichment of C. elegans ventral enclosure genes occurs with genes expressed during X. tropicalis neurulation, in which the neural tube is formed. This stage does not exist as such in nematodes where neurulation and muscle determination co-occur during ventral enclosure (Chin-Sang and Chisholm, 2000). Interestingly, we found that this overlap is strongly enriched for transcription factors for genes of the homeobox and LIM superclass (P<10-12 and P<10-6, respectively). Other groups of genes that are involved in neuronal specification are also enriched (P<10-8) in this set. It is interesting to speculate that the transcriptional correspondence between the ventral enclosure stage of the nematode species and the tail-bud and neurulation stages of X. tropicalis is a product of a diversification that occurred along the chordate lineage. A similar pattern was detected upon comparison with a recent Drosophila melanogaster timecourse (Graveley et al., 2011). The genes expressed at ventral enclosure are significantly enriched in D. melanogaster orthologs expressed during two disparate stages with a similar pattern of gene function enrichment found in the previous comparison. In both comparisons, C. elegans ventral enclosure genes are conserved across phyla in two distinct stages. This might suggest that the stage has subfunctionalized (Lynch and Force, 2000)into two distinct and temporally distant stages, or alternatively that two distinct ancestral milestones became coupled in C. elegans.
Our results expose the discontinuous nature of development through milestones – stages in which the similar morphology across species is paralleled by a conserved underlying molecular profile. We have provided evidence for this by examining species in which the timings of developmental markers have evolved yet the expression divergence between them is conserved. This “decoupling” criterion provides a simple operational definition of a bona fide stage – or milestone – in evolutionary terms and leads to the view that development is punctuated by milestones encompassing stable states of expression, bridged by considerable transcriptional turnover. Furthermore, together with our X.tropicalis and D. melanogaster comparison, we offer a model in which development differs across closely related organisms in the timing between and the details of conserved stages, while major phyla-level differences occur by the diversification of developmental milestones.
development evolution developmental biology evolution gene expression
")









1 Comment Leave a comment ›